




L’homme ne descend pas d’un primate arboricole ! Une évidence méconnue.
| L’homme ne descend pas d’un primate arboricole ! Une évidence méconnue. The ancestor of man was not an arboreal primate ! A disregarded evidence. Biom. Hum. etAnthropol. 1999, 17, p. 147-150. Y. Deloison. L’Homme primate arboricole ? Yvette Deloison |
Résumé
A la suite de l’étude de la spécialisation des extrémités des membres des Australopithèques, qui présentent des caractères d’arboricolisme, étant donné que l’évolution ne revient jamais en arrière (loi de Dollo), il apparaît évident que l’ancêtre commun des Australopithèques, des grands singes et de l’Homme était un Primate aux extrémités des membres indifférenciées. De plus, cet ancêtre devait être bipède ce que prouve, entre autres caractères, la structure primitive de la main humaine.
Mots-clés : bipédie -Australopithecus – main – pied.
Abstract
The foot bones of Australopithecines from Ethiopia and South Africa as well as ten fossil footprints of Laetoli have arboreal anatomical characters. The evolution never goes backward (law of Dollo), then it becomes obvious that the ancestor Australopithecines, apes and man had hand and foot whithout specialisation. The primitive hand of the man indures that his ancestor was neither arboreal nor quadrupedal consequently he was bipedal.
Keys words : bipedal locomotion -Australopithecus – hand – foot.
Le débat sur l’origine de la bipédie remonte loin dans le temps. En effet, nous sommes habitués aujourd’hui à l’idée que la bipédie humaine représente l’aboutissement d’une évolution qui nous a menés du tétrapode primitif à l’Homme érigé actuel. Le développement du cerveau est alors une conséquence de cette bipédie qui aurait libéré la main et entraîné l’augmentation de la capacité crânienne.
L’hypothèse des anthropologues, telle qu’elle fut développée entre autres par Howells et Wasburn (1953 et 1978) et qui continue de faire loi, considère que l’Homme a évolué à partir d’un primate arboricole et que son redressement se serait fait dans les arbres avant de descendre sur le sol. Autrement dit, que le pied de l’Homme actuel dériverait d’un pied de primate arboricole. Un des premiers à avoir émis cette idée est Huxley en 1868. Depuis, l’ensemble des anthropologues continue à raisonner sur ces a priori qui n’ont pas vraiment fait l’objet de démonstration scientifique.
Passée au crible de l’anatomie comparée, cette hypothèse ne résiste pas. Dans le principe, aucune hypothèse scientifique ne doit être conservée si elle n’est compatible avec les faits. En ce qui concerne l’origine humaine, ces faits sont d’ordres paléontologique et anatomique. Il est certain que tous les Primates, appartenant au même ordre, présentent un plan d’organisation commun où l’on retrouve : plantigradie, pentadactylie, mamelles pectorales, frontalité des orbites donnant une vision stéréoscopique etc… Mais, ce qui justifie la remise en cause des théories traditionnelles c’est: qu’elles n’expliquent pas les différences anatomiques que l’on observe entre l’Homme et les autres Primates.
Les Primates, comme d’autres animaux, se placent sur des voies que leur spécialisation a rendues irréversibles, que ce soit les tarsiers ou les simiens même anthropomorphes. Par exemple, le Tarsius a des membres postérieurs très longs adaptés au saut, les anthropomorphes (Gorille, Chimpanzé. Orang-outan) ont des membres adaptés à la suspension et à la prise des branches à la fois par les mains et les pieds. Il a une relation directe entre les proportions des membres et locomotion, les valeurs de l’indice intermembral des grands singes sont intéressantes à comparer à celles de l’Homme :

La main de l’homme est indifférenciée avec symétrie rayonnante et opposition du pouce aux autres doigts indifféremment et le pied, lui, est très spécialisé pour la bipédie permanente : adduction du premier orteil et prépondérante sur les autres orteils, présence d’une voûte plantaire avec relèvement antérieur du calcanéum. A l’inverse, les grands singes présentent des mains sans opposition du pouce mais avec un système musculo-tendineux facilitant la suspension, avec impossibilité d’étendre les doigts quand le membre antérieur est tendu et, entre autres caractères, des phalanges courbes. Cette courbure des phalanges existe aussi au pied qui a la faculté d’opposer le premier orteil aux orteils latéraux grâce à sa possibilité d’abduction de l’hallux.
Si l’Homme avait emprunté une des voies, dans lesquelles la main a un rôle dans la locomotion, il n’aurait jamais pu retourner à l’état initial indifférencié pour choisir un autre schéma morphologique, car la spécialisation anatomique ne régresse jamais : un cheval ne peut pas récupérer les doigts perdus au cours de son évolution, pas plus qu’un Orang-Outan ne verra repousser ses pouces atrophiés. La spécialisation peut s’arrêter à des stades différents mais ne revient jamais en arrière (Loi de Dollo). L’évolution ne connaît pas de repentirs.
Quels sont les stades de spécialisation de l’Homme ?
Chez l’Homme adulte actuel, la main a conservé une anatomie indifférenciée, c’est-à-dire non spécialisée ni pour la suspension, ni pour la locomotion, le pied, quant à lui, est hautement spécialisé dans la locomotion bipède de type humain. On peut même dire que la main est isotrope alors que le pied est anisotrope, c’est-à-dire qu’il n’a pas les mêmes propriétés suivant les mêmes directions (notons l’espace isotrope de la chondrification du carpe). Le pied de l’Homme ne possède aucune des caractéristiques de l’arboricolisme que l’on observe chez les grands singes, telles que la régression et l’abduction du premier orteil jointes à des orteils latéraux longs et fortement convexes.
Qu’en est-il chez les Primates fossiles ?
Laissons de côté les singes fossiles tels que Proconsul (20 millions d’années) ou Kenyapithecus déjà très engagés dans des voies de spécialisations simiennes et observons les Australopithèques. Nous constatons qu’ils possèdent, au niveau du pied, un ensemble de caractères arboricoles tels qu’on les observe chez les grands singes actuels : Gorille, Chimpanzé et Orang-Outan. Chez la célèbre Australopithèque Lucy, les caractères de bipédie se limitent au sacrum, mais on constate aussi une articulation tibiotalienne proche de celle de l’Orang-Outan dont on sait qu’il peut se tenir debout sur le sol avec un alignement total du corps et du membre inférieur. Cette dernière possibilité n’existe pas chez Pan et Gorilla. Ce qui laisse supposer cette possibilité chez Lucy, que l’on peut accorder aussi au dernier squelette, pratiquement complet, StW 573, découvert par Ron Clarke à Sterkfontein en Afrique du Sud en 1995. En effet, ce dernier possède le même type d’articulation tibio-talienne que Lucy c’est-à-dire proche de la forme de celle de l’Orang-Outan, à savoir les bords de la trochlée sont au même niveau, alors que chez les singes le bord médial présente un effondrement par rapport au bord latéral, sauf chez Pongo. Cependant, ces deux Australopithèques possèdent, en plus d’une articulation cunéométatarsienne du premier rayon trochoïdale, des phalanges longues et fortement incurvées, spécialisations typiques des membres arboricoles. L’ensemble des os du pied du squelette de Sterkfontein confirme par son anatomie la possibilité de préhensilité du pied et donc d’une locomotion arboricole. Ce pied n’a pu s’élaborer, comme tout organe spécialisé, qu’à partir d’un pied doté de toutes les potentialités. Le pied de l’ancêtre de l’Homme, quant à lui, a suivi un autre chemin de spécialisation. La forme qui donna naissance à ces différentes évolutions était forcément une forme indifférenciée. Dans la dernière publication de Ron Clarke (1999) portant sur l’observation des os de la main chez l’Australopithèque StW 573, il constate que les épiphyses du radius et de l’ulna ressemblent surtout à celles des grands singes, que les épiphyses distale de l’humérus et proximale de l’ulna ont des caractères proches de ceux de Pongo. Enfin, il en conclut que l’ensemble de ces caractères sont une indication de locomotion arboricole avec le membre supérieur.
Où chercher les origines de la forme ancestrale de la lignée humaine ?
Actuellement, on peut s’adresser à la paléontologie et à l’embryologie.
Jusques à quand doit-on remonter pour observer un pied indifférencié ? Nous n’en connaissons actuellement aucun fossile. Les plus anciens os de pieds fossiles d’Australopithèques ne datent environ que de trois à quatre millions d’années. Il faut donc remonter beaucoup plus loin. On constate que les premiers tétrapodes sortis des eaux primitives se sont tout de suite engagés dans des voies irréversibles, par exemple les amphibiens et les reptiles qui n’ont pu donner naissance à un pied du type primate.
L’ensemble des reptiles ayant subi par fusion une réduction. du nombre des os de la jambe et du pied au cours de l’évolution n’ont pu revenir en arrière et ne peuvent donc pas se situer dans la lignée des Primates, lesquels ont conservé l’ensemble des osselets tels qu’ils existaient chez les tout premiers mammaliens primitifs dès le Carbonifère. Ce phénomène de fusion d’os existe aussi chez les oiseaux qui peuvent, quant à eux, dériver des reptiles. La lignée des Mammifères pourrait à la rigueur avoir démarré à partir d’un modèle du genre Salamandre. Le squelette du pied et de la jambe de ce dernier genre présente une forme archaïque qui a pu évoluer vers divers modes de locomotion. Il s’agirait dans ce cas d’un amphibien primitif du type Urodéliforme (Westenhofer 1926). Chez les Mammifères, le calcanéum s’est déplacé vers la région plantaire du pied pour devenir sous-jacent au talus. Ce processus se retrouve au cours de la vie embryonnaire : au début, les deux os sont côte à côte, puis le calcanéum s’éloigne du talus qui demeure dans le prolongement du tibia. Le calcanéum accomplit ensuite un mouvement de rotation (supination) pour prendre sa place sous le talus en perdant son articulation avec la fibula.
La superposition du talus sur le calcanéum et les relations particulières des tibia et fibula avec le talus constituent cette cheville caractéristique des Mammifères. La formation de la cheville est de toute évidence la conséquence du redressement du mammalien primitif. Au cours de ce redressement, l’os central (naviculaire des Mammifères) s’est déplacé vers le bord du pied, et ensuite, et de manière concomitante, il s’est produit une pronation du pied amenant l’ensemble de la plante sur le sol depuis le bord latéral, permettant ainsi une meilleure stabilité, condition de la plantigradie .
Rappelons que tous les Mammifères plantigrades ont le basipode (calcanéum) relevé vers l’arrière sauf l’Homme qui l’a relevé vers l’avant.
Ne nous étonnons pas qu’il existe si peu de fossiles ; il est certain que la fossilisation très ancienne d’organismes graciles n’a pu se produire que dans des conditions exceptionnelles. Or, si l’ancêtre de l’Homme était déjà un vertébré terrestre, les chances de fossilisation à l’air libre étaient très réduites. On trouve surtout des espèces aquatiques, poissons, coquillages, ou bien terrestres susceptibles d’enlisement. Il faut être conscient que le nombre de fossiles est très réduit et même dérisoire par rapport au nombre réel d’animaux qui ont existé.
L’embryologie peut-elle nous apporter des éclaircissements ?
A un certain stade de la vie embryonnaire, la morphologie est semblable entre des embryons d’animaux aussi différents que l’Homme, le Poulet, le Chien ou la Tortue (Haeckel 1868).
Chez l’Homme, la première ébauche du membre supérieur apparaît dès le 24e jour après la conception, celle du membre inférieur vers le 26e jour, donc très tôt. Avant leur forme définitive, la main et le pied présentent une forme de palette dans laquelle très vite on distingue les cinq rayons. La main gardera sa forme rayonnée n’abandonnant que la palmure. A 49 jours, le pied de l’embryon humain a déjà sa forme définitive de pied d’Homme adulte. Sur des coupes de pieds d’embryons humains de 43 jours, on voit déjà très bien la forme du calcanéum. L’ensemble des os du tarse et du métatarse sont déjà en place. On mesure l’importance de cette constatation quand on sait que la différenciation sexuelle n’est pas encore effectuée. Lecalcanéum présente dès la fin de la vie foetale des caractères spécifiquement humains, par exemple, le tubercule externe qui ne peut pas être une conséquence de la locomotion bipède permanente puisqu’il précède la vie extra-utérine.
En étudiant l’embryologie du pied, on ne peut s’empêcher d’être surpris par la précocité de la formation de cette spécialisation exclusivement humaine.
Résumons donc. L’ancêtre commun des grands singes et de l’Homme, forcément très ancien, avait un pied non spécialisé possédant des potentialités variées d’évolution.
Cet ancêtre commun, qu’était-il ? Examinons plusieurs hypothèses :
– arboricole ? – Non, car il aurait acquis des spécialisations irréversibles comme nous l’avons dit plus haut notamment au niveau de la main. Cet ancêtre devait garder ouvert l’éventail des possibilités.
– terrestre ? – Dans ce cas, était-il quadrupède ou bipède ?
Dans le cas d’un ancêtre commun quadrupède, au niveau des petits singes, cette hypothèse est compatible avec leur mode de locomotion. En effet, ils marchent avec les mains à plat sel-le support alors que les grands singes reposent sur le dos des deuxièmes phalanges des mains, les pieds restant à plat sur le sol comme chez les autres Primates. La quadrupédie des grands singes étant un peu particulière, nous y reviendrons.
Quant à l’Homme, son ancêtre n’a pas pu être quadrupède sinon sa main aurait évolué et ne serait pas restée archaïque sans ébauche même de spécialisation. Elle n’a donc servi ni d’appui ni de moyen de préhension. Le squelette du pied de l’Homme ne repose pas à plat sur le sol comme chez les autres Primates, mais forme une voûte plantaire qui ajoute à la solidité de son pied appelé à supporter la totalité du poids du corps. De plus, le poids du corps est amorti au niveau de !a plante du pied par une épaisse couche de graisse chez l’Homme, alors que la sole plantaire du pied des grands singes en est dépourvue.
Toutes ces données semblent orienter vers un ancêtre commun non quadrupède et donc bipède, mais d’un type de bipédie primitive différente de celle de l’Homme actuel.
Ainsi le fait que l’ancêtre commun ait été bipède pourrait-il expliquer que la main soit restée primitive et donc non spécialisée. Au cours du temps, cette bipédie s’est affirmée par adaptation fonctionnelle avec la pratique régulière de la marche bipède conduisant à la formation anisotrope des os du pied. avec le redressement de la partie antérieure du calcanéum et :a formation de la voûte plantaire chez [Homme. Cette dernière représente un trait anatomique de l’Homme, unique chez les Primates.
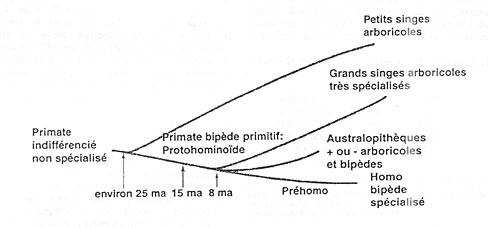
Les données fournies par les généticiens semblent apporter une confirmation à nos observations. Ils avancent d chiffres qui sont compatibles avec les idées exprimées plus haut, à savoir, le tronc commun aux Primates se situerait vers 4(.t millions d’années, les petits singes se séparant des autres Primates vers 25 millions d’années.
L’ancêtre commun aux grands singes et à l’Homme, que nous nommons Protohominoïde, se situerait vers 15 millions d’années et serait aussi à l’origine des Australopithèques. Ces derniers ainsi que les grands singes se seraient séparés de cette lignée vers 8 millions d’années. Quand les Gorilles ou les Chimpanzés se déplacent sur le sol, Es sont en position que l’on peut qualifier de mi-bipède mi-quadrupède. Cette attitude, c’est-à-dire la position des maires sur le sol reposant sur le dos des deuxièmes phalanges, et non la main à plat comme les petits singes,
Cette adaptation à une telle forme de quadrupédie et à l’arboricolisme expliquerait la grande spécialisation de leurs membres. Remarquons que c’est durant sa petite enfance que le Chimpanzé ressemble le plus à l’humain, sa tête est droite sur la colonne, c’est en devenant adulte que les différences avec l’Homme vont en s’accentuant.
Les traits résiduels de bipédie observés au sacrum des Australopithèques ainsi que les caractères de leurs membres spécifiques de la locomotion arboricole pourraient s’expliquer par une adaptation à une nouvelle forme de vie incluant le grimper dans les arbres. Ayant disparu il y a au moins un million d’années, ils n’ont peut-être pas eu le temps de parfaire leur spécialisation comme l’ont fait les grands singes qui existent toujours à l’heure actuelle.
Le schéma suivant illustre cette ancestralité de la bipédie.
Signalons que parmi les Mammaliens, ce sont les Primates qui possèdent la structure osseuse du pied la plus primitive et parmi les Primates, ce sont les Hommes qui présentent un maximum de caractères primitifs, vestiges de l’adaptation à une lointaine vie aquatique que l’on retrouve dans la construction d’organes tels que la rate ou le rein ou encore l’appareil digestif.
Le simple et trop rapide énoncé de réalités contrôlables et de constatations avérées avive encore davantage l’intérêt que soulève depuis longtemps la question primordiale de la bipédie humaine. Ces données offrent une nouvelle base de réflexion à tous ceux que préoccupe la recherche fascinante des origines de l’Homme.
Bibliographie
CLARKE R.J. 1999 — Discovery of complete arm and hand of the 3.3 millionyear-old Australopithecus skeleton from Sterkfontein. South.African Journal of Sciences, 95, nov./déc. 1999. P. 477-480.
HOWELLS W. Préhistoire et histoire naurelle de l’Homme. Payot, 1953, 331p.
WASHBURN S. L’évolution de l’Homme in l’Evolution. Bibi. Pour la Science, Belin, 1978, p. 98-107.
WESTENHOFER Prof. Dr M. 1926 — Vergleichend-morphologische Betrachtungen über die Entstehung der Ferse und des Sprunggelenks der Landwirheltiere mit besonderer Beziehung auf den Menschen. Archiv. Für Frauenkunde und Konstitutions Forschung. 12, 1-48, Leipzig.
Contact : Yvette Deloison,
Chargé de Recherche au CNRS, EP 1781 CNRS. 44 l’Amiral-Mouchez 75014 Paris.
Article reçu le 9/10/99, accepté le 15/11/99
Les liens présentés dans cette page sont choisis par Hominides.com
Du même auteur, Yvette Deloison  | Préhistoire du piéton Et si vous changiez d’avis sur la bipédie humaine ? Yvette Deloison développe sa théorie de la bipédie originelle. L’homme n’aurait pas acquis la marche bipède mais il l’aurait au contraire conservée… contrairement aux grands singes. Ce livre bouscule les idées reçues mais son argumentation et les recherches présentées forcent à la réflexion… |
A lire également :
La bipédie chez l’homme
Les caractéristiques de la bipédie identifiée sur un fossile
Les traces de pas à Laétoli
2019 Une nouvelle espèce de grand singe bipède… dans les arbres !
2022 Toumaï, la bipédie humaine il y a 7 millions d’années









Sous la direction de Jean-Jacques Hublin


Bertrand Roussel



